ecomorphology & evolution of the avian sternum
|
The evolution of powered flight in early birds and their dinosaur ancestors was a key locomotor innovation that expanded the ecological potential of basal birds and greatly contributed to their remarkable diversity today. Despite a rich body of literature on the origin of flight, details of how flight first originated are contentious, and opinions are primarily split between the Trees-down (gliding first) hypothesis, and the ground up (running first) hypothesis. Most research on the origin of flight has focused on the morphology of the wings, and has largely ignored other important elements of the skeletal system. My primary dissertation research focuses on the ecological significance of the morphology of the sternum -- or breastbone, which sits in the middle front portion of the ribcage. Modern flying birds have greatly enlarged sterna, with a median bony projection, or keel, that serves as the attachment site for the major flight muscles that originate from the wings. Since this element is so crucial to flight in living birds, a robust analysis of the relationship between sternum morphology and flight ability is long overdue. To test this, I am investigating which aspects of sternum morphology are associated with different locomotory abilities (ie. wing-propelled diving, thermal soaring, continuous flapping, flightlessness) through a combination of 3D landmark-based geometric morphometrics and phylogenetic comparative methods, including distance-based phylogenetic generalized least squares regressions. Understanding this relationship in living bird species will allow me to make inferences on the possible locomotion and flight abilities in fossil birds, which will further our understanding of the evolution of flight across the dinosaur-bird transition.
The extant portion of this project is now published in BMC Biology. |

3D model of of Anhinga anhinga sternum with landmarks, used in geometric morphometric analysis. Screenshot taken on 3D imaging software Avizo.

Sternum in fossil specimen of enantiornithine bird Longirostravis hani from the Late Cretaceous of China. Photo taken at the Institute of Vertebrate Palaeontology and Palaeoanthropology in Beijing, China.
|
Phylogenetic affinities of of early european passeriformes
|
Passeriformes, colloquially known as perching birds, is the most taxonomically diverse and globally widespread order of living birds, consisting of two main suborders, oscines and suboscines. Phylogenetic evidence suggests that Passeriformes evolved from the fossil group Zygodactylidae, which shares a similar zygodactyl foot morphology with the closest living relatives to passerines, the Psittaciformes (parrots). Three well-preserved passerine-like fossils with ambiguous phylogentic positions are known from the early Eocene of Europe: Resoviaornis jamrozi, Wieslochia weissi, and Jamna szybiaki. The goal of this project is to perform the first set of phylogenetic analyses that incorporates all three of these fossils, and the first cladistic analysis on the fossil bird Resoviaornis. Using a combination of morphological characters and DNA data, we performed phylogenetic analyses using maximum parsimony, maximum likelihood, and Bayesian inference hypotheses. Resoviaornis consistently grouped with oscines, while Wieslochia falls within suboscines, a result that is consistent with previous hypotheses. The affinities of Jamna were not as well-resolved, likely due to the fossil missing a lot of key features, however with some analyses placed it within oscines and others in the stem of extant Passeriformes. Resolving the phylogenetic relationships of these early Passeriformes can not only provide temporal information for calibrating molecular phylogenies, but also insight in the early radiation and biogeography of the most diverse group of birds.
|

Slab and counterslab of Resoviaornis jamrozi from the early Eocene of Poland. Photos courtesy of Zbigniew Bochenski and Krzysztof Wertz.

Slab specimen of Jamna szybiaki from the early Eocene of Poland. Photos courtesy of Zbigniew Bochenski and Krzysztof Wertz.
|
Cranial variation in hadrosaurines from the Dinosaur Park formation in Alberta, Canada
|
The Dinosaur Park Formation of Alberta documents one of the most diverse assemblages of hadrosaurine dinosaurs. Historically, two species of the genus Gryposaurus have been recognized in the Dinosaur Park Formation, G. notabilis and G. incurvimanus, which are differentiated primarily on their nasal arch morphology. These two species have recently been suggested to represent either variable morphs within the species G. notabilis (e.g., representing different ontogenetic, or life stages) or two distinct taxa within an evolving Gryposaurus lineage (e.g., anagenesis). These alternative hypotheses have never been adequately tested via detailed morphological comparisons, morphometrics, or biostratigraphy. For my undergraduate thesis project, I performed a geometric morphometric analysis of hadrosaurine skulls from the Dinosaur Park Formation to assess the influence of ontogeny on skull morphology. G. incurvimanus skulls were found to be distinctly smaller, and morphologically different from those classified as G. notabilis, with the larger G. notabilis skulls having higher nasal arches that are set farther back on the skull, a feature commonly seen in adult individuals of other hadrosaurids, such as Brachylophosaurus and lambeosaurines. High resolution stratigraphic data were used to map this morphology through time to evaluate the anagenesis hypothesis. The stratigraphic distributions of the two species showed considerable temporal overlap, which indicates that the sampled individuals lived over a relatively short period of time (< 0.5 million years). Overall, our results suggest that G. incurvimanus and G. notabilis may represent two different life stages within one species, G. notabilis, with specimens previously classified as 'G. incurvimanus' actually representing the juvenile individuals. This study improves our understanding of the extent of potential individual variation within a single Gryposaurus species, which can be useful in assessing the validity of other hadrosaurine species. This study has been published in the Canadian Journal of Earth Sciences.
|
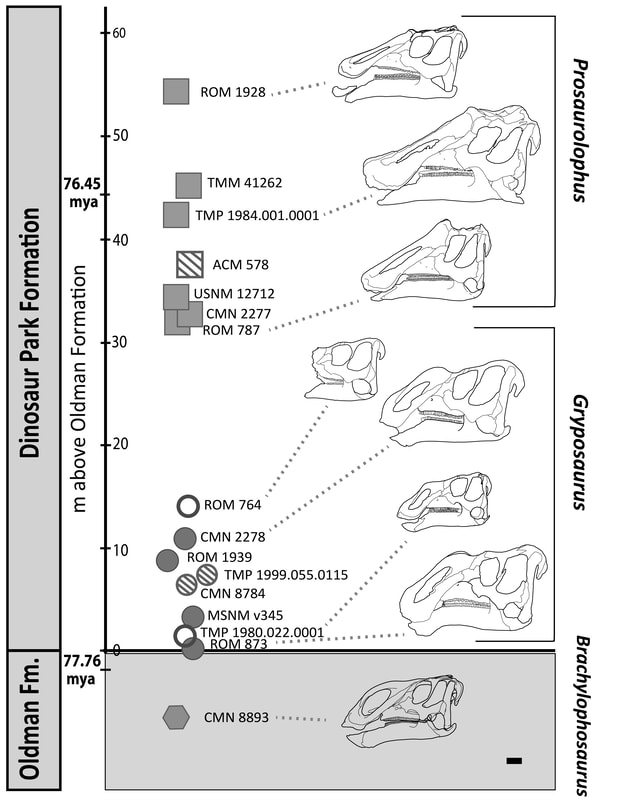
Figure from Lowi-Merri and Evans (2020). Temporal range of hadrosaurines from Dinosaur Provincial Park; Gryposaurus and Prosaurolophus specimens from the Dinosaur Park Formation with known stratigraphic data are included. The hexagon indicates Brachylophosaurus. Closed circles indicate specimens previously identified as G. notabilis, open circles indicate those previously identified as ‘G. incurvimanus’, and striped circles indicate Gryposaurus sp. Closed squares indicate Prosaurolophus maximus, Skull illustrations are scaled against each other to show size variation. Stratigraphic dates sourced from Freedman Fowler and Horner (2015). Scale bar: 10 cm.
|
